|
アミノ基転移 グルコース-アラニン回路 酸化的脱アミノ その他の脱アミノ アミノ酸の脱炭酸 アミノ酸骨格の代謝 アミノ酸代謝の臓器特異性 アミノ酸代謝酵素の欠損症 |
  |
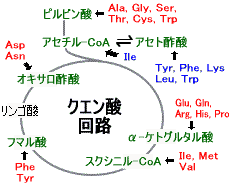
脱アミノ化されて生じるα-ケト酸は下の図のような経路で,(1) 糖の合成,(2) ケトン体や脂肪酸の合成に利用される。アミノ酸によっては,(1) と(2)の両方に関わるものもある。
青はケト原性アミノ酸,赤は糖原性アミノ酸
|
|
|
  |
|
|
|
|
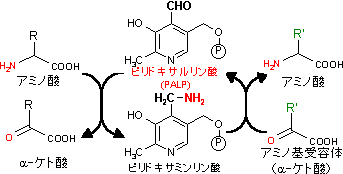
現在,アミノトランスフェラーゼとして基質特異性を異にする50種以上の酵素が知られている。いくつかの例を次に示す。
Asp + a-ケトグルタル酸 オキサロ酢酸 + Glu アスパラギン酸アミノトランスフェラーゼ (AST) Ala + a-ケトグルタル酸 ピルビン酸 + Glu アラニンアミノトランスフェラーゼ (ALT) Tyr + a-ケトグルタル酸 4-ヒドロキシフェニルピルビン酸 チロシンアミノトランスフェラーゼ Leu + a-ケトグルタル酸 a-ケトイソカプロン酸 + Glu 分枝鎖アミノ酸アミノトランスフェラーゼ Glu + グリオキシル酸 a-ケトグルタル酸 + Gly グルタミン酸アミノトランスフェラーゼ Ala + グリオキシル酸 ピルビン酸 + Gly アラニン-グリオキシル酸アミノトランスフェラーゼ Ser + ピルビン酸 ヒドロキシピルビン酸 + Ala セリン-ピルビン酸アミノトランスフェラーゼ g-アミノ酪酸 + a-ケトグルタル酸 コハク酸セミアルデヒド + Glu g-アミノ酪酸アミノトランスフェラーゼ Orn + a-ケトグルタル酸 グルタミン酸セミアルデヒド + Glu オルニチンアミノトランスフェラーゼ
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
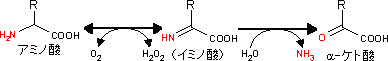
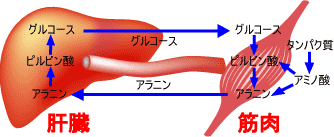
全てのアミノ酸のアミノ基はアスパラギン酸とグルタミン酸に集められ,ミトコンドリア内に送り込まれる(アスパラギン酸とグルタミン酸はミトコンドリア内膜を通れる)。
グルタミン酸は,ミトコンドリアにおいて,グルタミン酸デヒドロゲナーゼによって酸化的に脱アミノ化され,α-ケトグルタル酸になる。これを酸化的脱アミノ(Oxidative
deamination)という。グルタミン酸デヒドロゲナーゼは補酵素としてNAD+を用いるが,生物によってはNAD+とNADP+の両方が用いられる場合もある(珍しい例)。
α-ケトグルタル酸はTCA回路の一員である。遊離したNH3は尿素回路によって直ちに尿素に変換される。
|
|
|
|
|
|
|
|
[L-アミノ酸オキシダーゼとD-アミノ酸オキシダーゼ]
肝臓,腎臓のペルオキシソーム中に存在する酵素で,FADやFMNなどが補酵素として結合している。アミノ酸に対する特異性は低く,Ser, Thr,
His, Gly, Glnなどが脱アミノ化される。反応の途中で生じるFADH2はO2で酸化され,過酸化水素 H2O2を生成する。
FADH2 + O2 ![]() FAD + H2O2
FAD + H2O2
図の2段階目の反応は非酵素的に進行する。
生体内には少なからずD-アミノ酸が存在する。特に,D-アスパラギン酸とD-セリンは比較的多量に存在する。D-アミノ酸は種々の臓器で発生の早い時期に一過性に増加する。D-セリンは中枢神経の興奮性アミノ酸受容体のアロステリック作動薬として作用することが知られている。D-アミノ酸はラセマーゼによってL-アミノ酸に変えられて代謝される以外に,上の図のように,D-アミノ酸オキシダーゼで酸化的脱アミノを受ける。
|
|
|
|
|
|
|
|
アミノ酸は脱炭酸により一級アミンを生じる。動物でもこれらの反応は見られるが,微生物では特に発達している。反応にはPALPを必要とする場合が多い。生成するアミンは強い生理活性をもつものが多く,”生理活性アミン(biogenic
amine)”と呼ばれる。
アドレナリン,ノルアドレナリン,ドーパミン,g-アミノ酪酸 (GABA),セロトニン,ヒスタミンなどはアミノ酸由来のホルモン,神経伝達物質である。
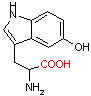 |
ヒドロキシ トリプトファン デカルボキシラーゼ |
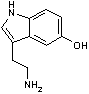 |
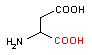 |
アスパラギン酸-1- デカルボキシラーゼ |
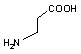 |
| 5-ヒドロキシトリプトファン | CO2 | セロトニン | Asp | CO2 | b-アラニン |
 |
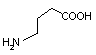
オルニチン デカルボキシラーゼ |
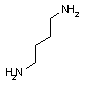 |
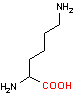 |
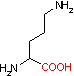
リシン デカルボキシラーゼ |
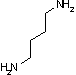 |
| Orn | CO2 | プトレッシン | Lys | CO2 | カダベリン |
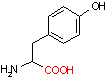 |
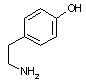
チロシン デカルボキシラーゼ |
 |
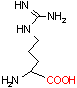 |
アルギニン デカルボキシラーゼ |
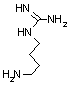 |
| Tyr | CO2 | チラミン | Arg | CO2 | アグマチン |
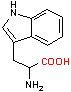 |
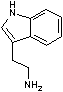
トリプトファン デカルボキシラーゼ |
 |
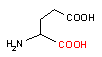 |
グルタミン酸 デカルボキシラーゼ |
 |
| Trp | CO2 | トリプタミン | Glu | CO2 | g-アミノ酪酸 |
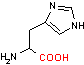 |
ヒスチジン デカルボキシラーゼ |
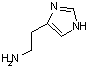 |
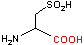 |
システイン スルフィニル酸 デカルボキシラーゼ |
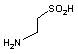 |
| His | CO2 | ヒスタミン | システインスルフィニル酸 | CO2 | ヒポタウリン |
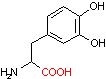 |
DOPA デカルボキシラーゼ |
 |
ドーパミン-b- モノオキシゲナーゼ |
 |
フェニルエタノール アミン N-メチル トランスフェラーゼ |
 |
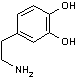
| 3,4-ジヒドロキシフェ ニルアラニン (DOPA) |
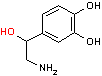
CO2 | ドーパミン | O2 H2O AsA DHAsA |
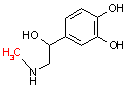
ノルアドレナリン | SAM S-AH | アドレナリン |
| AsA: アスコルビン酸 (ビタミンC) DHAsA: デヒドロアスコルビン酸 |
SAM: S-アデノシルメチオニン S-AH: S-アデノシルホモシステイン |
|||||
|
|
|
|
|
|
|
|
| オキサロ酢酸群 ピルビン酸群 α-ケトグルタル酸群 スクシニル-CoA群 アセチル-CoA群 フマル酸群 |
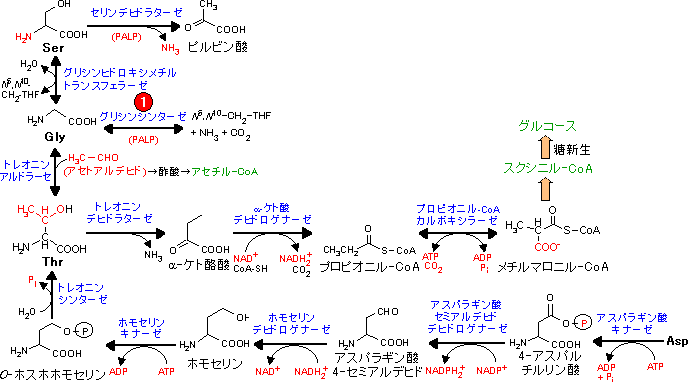
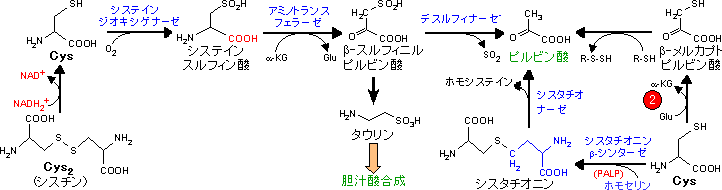
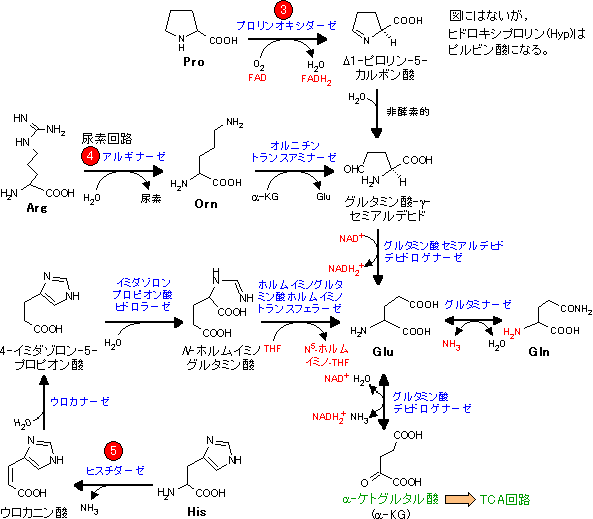
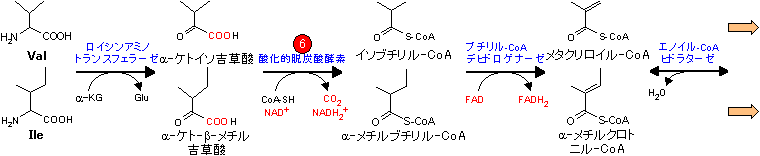
アミノ酸骨格の代謝は個々のアミノ酸で異なり,分解と合成は互いに関連している。
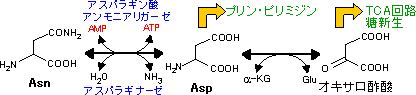
 Asp, Asnの代謝 (オキサロ酢酸群) Asp, Asnの代謝 (オキサロ酢酸群) |
| Ala, Gly, Ser, Thr, Cysの代謝 (ピルビン酸群) |
筋肉のタンパク質分解によって生じたアラニンは肝臓でピルビン酸に戻され,グルコース合成に利用される。
| Pro, Arg, His, Glu, Glnの代謝 (α-ケトグルタル酸群) |
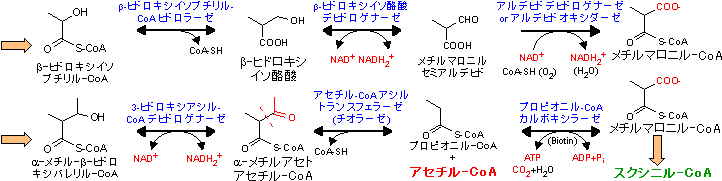
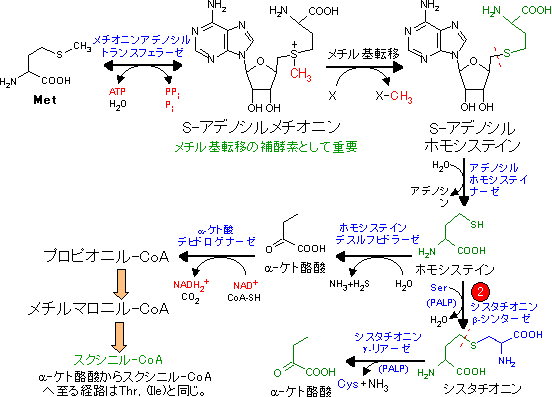
| Val, Ile, Metの代謝 (スクシニル-CoA群) |
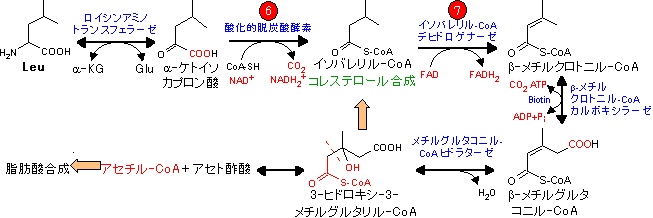
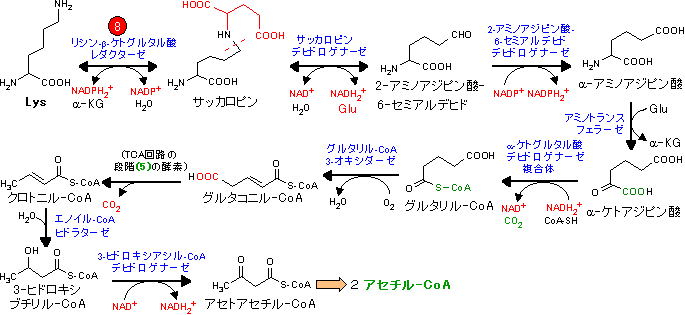
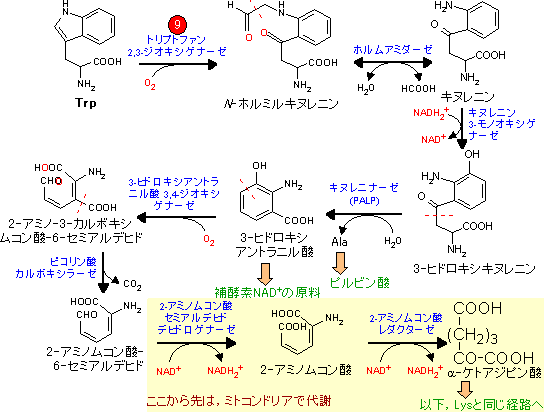
| Leu, Lys, Trpの代謝 (アセチル-CoA群) |
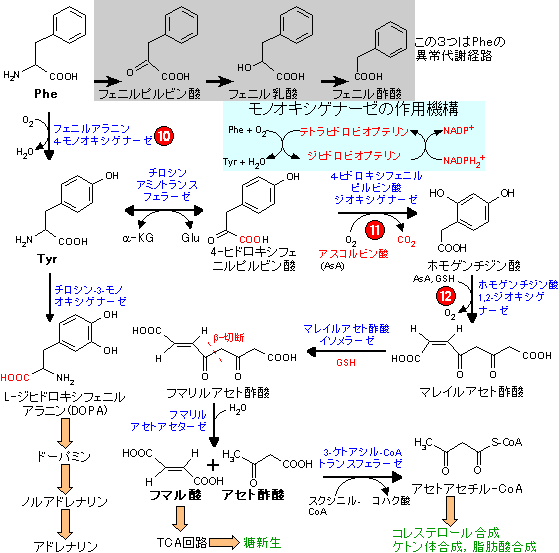
| Tyr, Pheの代謝 (フマル酸群) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
酵素 疾病 症状 高グリシン血症 脳脊髄液や尿中のグリシン濃度が上昇。重症の精神身体発育阻害,発作がみられ,小児期に死亡。 シスタチオニン-β-シンターゼ欠損症 血中メチオニンの上昇とホモシスチン尿症がみられる。精神発達障害,水晶体転位,骨格異常など。 高プロリン血症 血中プロリン濃度が上昇。 高アルギニン血症 尿素回路を参照 ヒスチジン血症 血中ヒスチジンの上昇。発育障害を起こす。精神発達障害や言語障害がみられることもある。 メープルシロップ尿症 尿がメープル(楓)シロップ様の匂いを発し,低分岐アミノ酸食治療をしないと,生後間もなく死亡する。 イソ吉草酸血症 ロイシンの中間代謝物が蓄積する。 高リシン血症 血中リシン濃度が上昇。 先天性トリプトファン尿症 尿中トリプトファン濃度の上昇。 フェニルケトン尿症 チロシンへの変換が行えないため,血中のフェニルアラニン濃度が上昇し,尿にフェニルピルビン酸が排泄される。知能低下,メラニン色素欠乏などを引き起こす。 チロシン症 尿に4-ヒドロキシフェニルピルビン酸が排泄される。肝臓や腎臓の不全を引き起こす。 アルカプトン尿症 尿に大量のホモゲンチジン酸が排泄され,空気酸化で黒変する。症状は晩年に関節炎がでる程度。
|
|