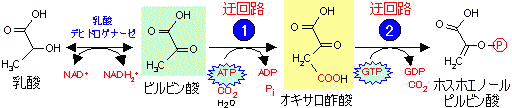
乳酸、ピルビン酸、
アミノ酸、プロピオン酸などから、おおむね解糖を逆行してD-グルコースをつくる経路を
糖新生(Gluconeogenesis)という。脂肪酸やアセチルCoAはピルビン酸に変換できないので,この代謝経路にのらない。ピルビン酸からグルコースに至る全反応は次のようになる。
2 Pyruvate + 4 ATP + 2 GTP + 2 NADH2+ + 6 H2O → Glucose + 4 ADP + 2 GDP + 6 Pi + 2 NAD+
段階
(1)(5)でATP,
(2)でGTPが消費される。この経路の最終段階の酵素(
グルコース-6-ホスファターゼ)は肝臓と腎臓にしか存在しない(「肝腎かなめ」という)。従って,糖新生でグルコースをつくるのは肝臓で行われ,他の臓器ではグルコール
6-リン酸までである。
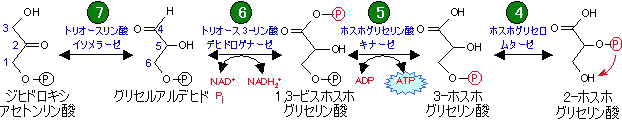
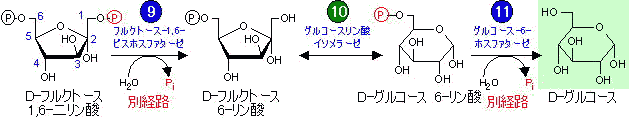
解糖の1、3、10番目の段階(糖新生では段階
(1)(2)、
(9)、
(11)に相当)は
不可逆であるため、これらの段階は別の経路または別の種類の反応が利用される(
「異化と同化は別経路」の例)。これによって、一見単なる逆反応のように見える糖新生と解糖を独立に制御できる。糖新生で見過ごせない事は,段階
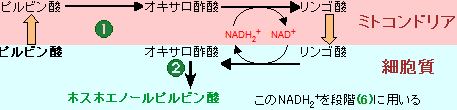
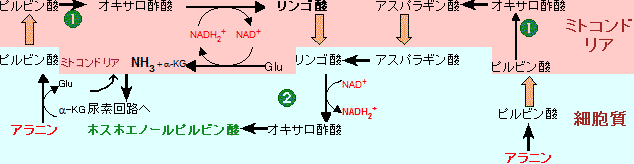
(6)でNADH
2+を必要とする点である。解糖や糖新生に利用できるNADH
2+の量は限られている。NADH
2+は
ホスホグルコン酸回路から60%,
リンゴ酸から40%が供給される。


は解糖の反応
の逆とは異なる経路
と
は解糖の反応
と
の逆とは異なり,加水分解。 酵素

脱アミノ
アセチルCoA