|
|
 
|
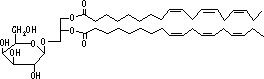
|
|
|
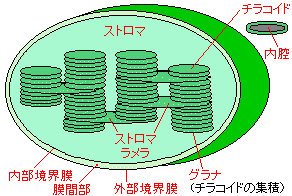
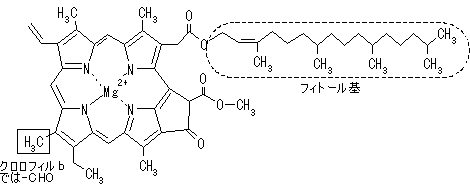 |
 |
||
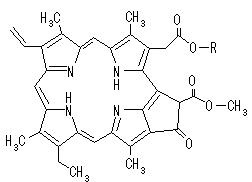
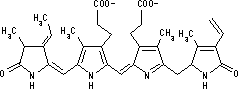
| [クロロフィル a (Chl a)の構造] | [フェオフィチンa (Pheo a)の構造] | ||
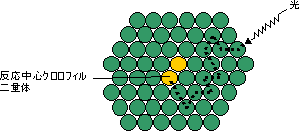
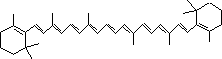
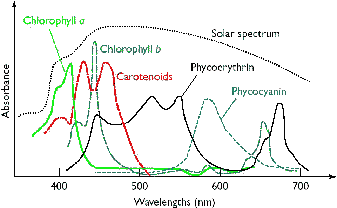
| クロロフィルはプロトポルフィリン IXの誘導体で,中心にMg2+が配位している。クロロプラスト中の大部分のクロロフィルは光を集めるアンテナの役割を果たす。吸収された光子のエネルギーはアンテナクロロフィル間を励起エネルギーとして移動し,アンテナクロロフィルよりも励起エネルギーの低い反応中心クロロフィルに集められる。クロロフィルが吸収できない波長の光を集めるために,カロテノイド(橙色),フィコエリトロビリン(赤色),フィコシアノビリン(青色)など,別の色の色素も使われる。 バクテリアのクロロフィルは,フィトール基の代りにゲラニルゲラニル基を持つものもある。また,ポルフィリン環の側鎖にも違いがある。 |
|
 |
 |
| [アンテナクロロフィルから反応中心クロロフィルへの エネルギーの移動] |
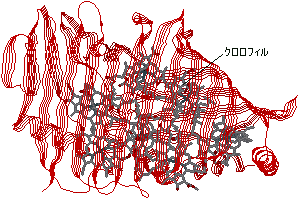
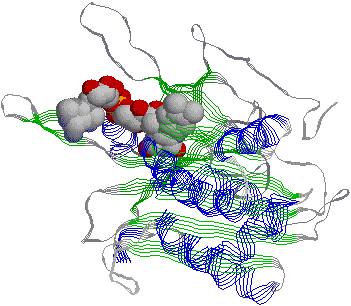
[光合成細菌のアンテナクロロフィルタンパク質] |
| 光合成細菌 (P. aestuarii)のバクテリオクロロフィルaタンパク質 7つのクロロフィルa分子を含んでいる。 |
 |
 |
| [b-カロテン] | [フィコエリトロビリン] |
 |
 |
| シアノバクテリア(C. caldarium)のフィコシアニンの構造] タンパク質(2量体)に3分子のフィコシアノビリンが結合している。 |
[クロロフィルa,b他の吸収スペクトル] |
|
|
 明反応の光化学系(PS)複合体
明反応の光化学系(PS)複合体| 複合体 | 成分名 |
|---|---|
| 光化学系(PS) 複合体II |
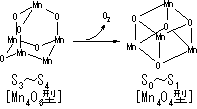
酸素発生複合体(水デヒドロゲナーゼ, OEC), P680, フェオフィチン(Pheo), 膜結合型プラストキノン(QA, QB), 膜結合型プラストキノンプール |
| シトクロム b6-f複合体 |
シトクロムb6(2ヘム型), (2Fe-2S), シトクロムf, 膜結合型プラストキノール(PQ) |
| プラストシアニン | プラストシアニン(シトクロム c-553, 10.5kD, CuI/II型) |
| 光化学系(PS) 複合体I |
P700, A0, A1 X,A,B(膜結合4Fe-4S型タンパク質), フェレドキシン, フェレドキシン-NADP+ レダクターゼ |
| 光リン酸化系 | ATP シンターゼ(ATPase) CF0:プロトン輸送チャネルタンパク質他 CF1:a3b3gde |
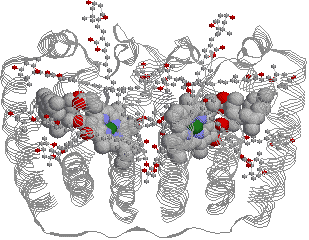 |
| [A. carteraeの集光性複合体(LHC)] |
| 中央にクロロフィルaが2個あり,その周りにたくさんのカロテノイド(peridinin)が存在する。 カロテノイドが集めた光はクロロフィルに渡され,次いで,P680やP700へ送られる。 |
植物光合成の光化学系(Z機構)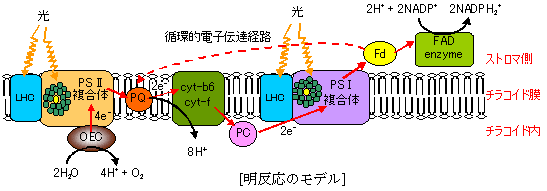 |
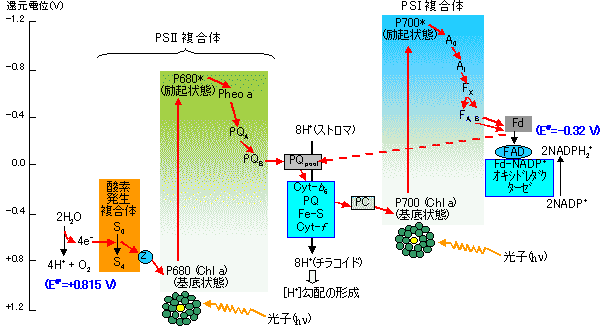 |
| [明反応のZ機構] |
 |
 |
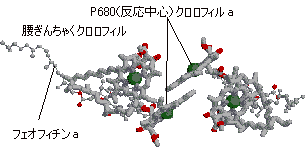 [色素だけを上から見た眺めた図] |
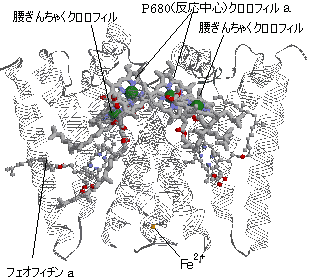
| [ほうれん草光化学系IIの反応中心の様子] P680タンパク質は四量体を形成。中央上部にP680クロロフィルa二量体(反応中心)が見える。 ポルフィリン環の面を向け合ってクロロフィル二量体になる。 腰ぎんちゃくクロロフィルを介してフェオフィリンaに電子が渡される。 |
|
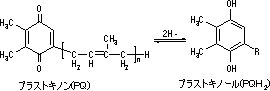
 |
 |
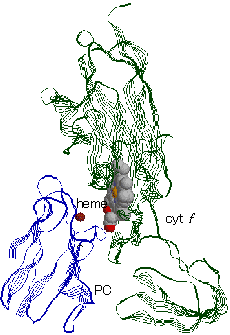 [シトクロムf-プラストシアニン複合体] 左下に,銅イオンを結合したプラストシアニン,右に変形型ヘムcをもつシトクロムfが見える。 |
|
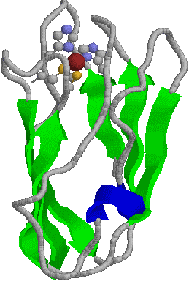
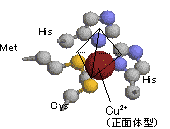
| [プラストシアニンの構造] | |||
| プラストシアニンはチラコイド膜の内表面に存在する10.5 kDaの青色銅タンパク質である。触媒部位のCu2+には,Cys,Metおよび2つのHisが配位している。 Cu2+ + e- プラストシアニンのCu2+は正方形型(E0'=+0.158 V)ではなくて正四面体型(E0'=+0.370 V)であり,大変,電子を受け取り易くなっている。 |
|||
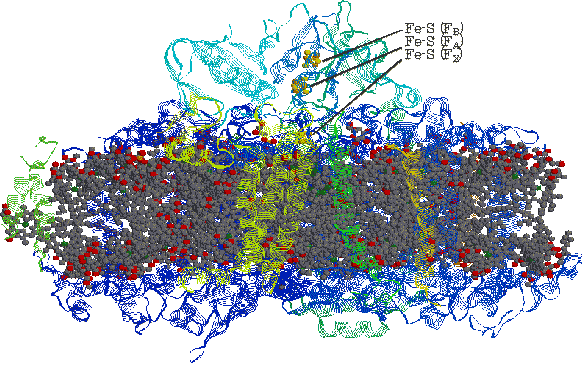 |
| [細菌光化学系複合体Iの立体構造(膜面から見た図)] |
| 色素分子は濃灰色で示す。2つの大サブユニット,9個の小サブユニット,[Fe-S]で構成されている。 95分子のクロロフィルa,22分子のb-カロテン,2分子のフィロキノンが含まれる。 |
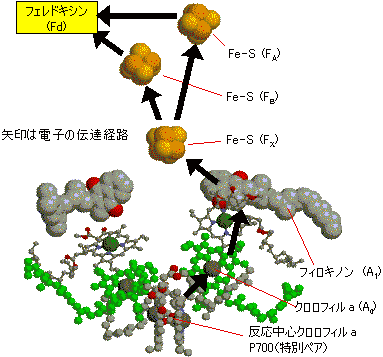 |
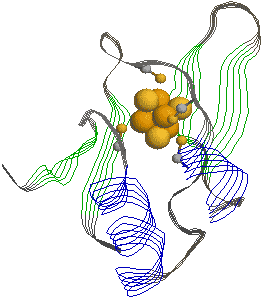
| [光化学系複合体Iの活性中心の様子] |
 |
 |
| [フェレドキシンの構造] | [フェレドキシン-NADP+ レダクターゼ] |
| フェレドキシンはストロマにある可溶性の酵素で, 4Fe-4S型の鉄-硫黄クラスターをもつ。 一度に1個の電子を受け取る。 |
レダクターゼ(緑)はFAD(図左上)を補酵素にもち, NADP+ ,2H+ ,4e-からNADPH2+をつくる。 フェレドキシンは青で示してある。 |
|
|
紅色細菌の光化学系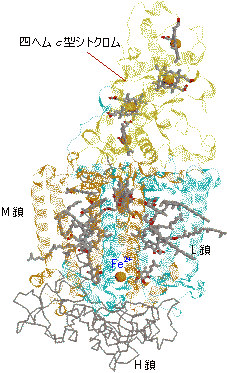 |
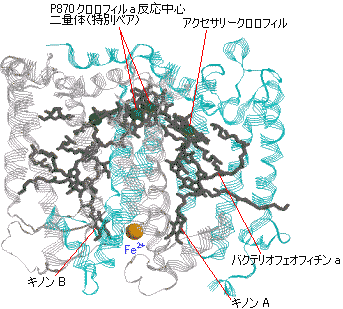 |
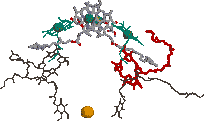
| [紅色細菌Rb. viridisの光化学系複合体] | [紅色細菌の光化学系の活性中心] |
| P870*(BChr a二量体) | BPheo a | |
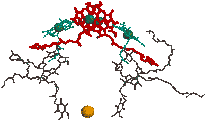 |
 |
|
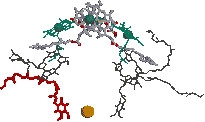
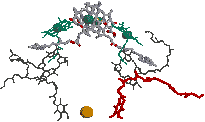
| 0 s (光照射直後) | 3 ps |
|
 |
 |
|
| キノンB (QB) 0.1 ms | 200 ps キノンA (QA) | |
| [紅色細菌の光化学系における電子伝達の様子] 励起された分子を赤で示す。 |
||
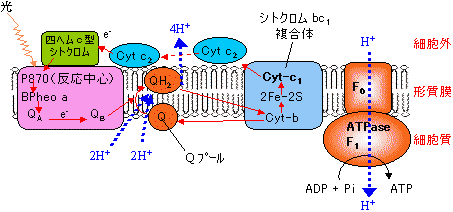 |
| [紅色細菌の明反応とATP合成] |
高度高塩菌の光合成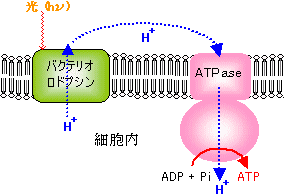 |
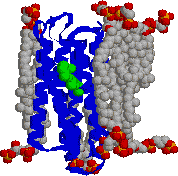 |
| [高度高塩菌の光合成機構] | [バクテリオロドプシンの立体構造] |
| りん脂質二重層を7本のへリックスで貫通した タンパク質がバクテリオロドプシンで,中央に レチナール分子(緑色)が見える。 |
|
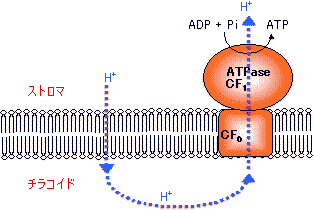 |
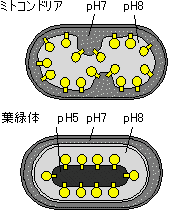 |
| [H+輸送ATPaseによるATP合成] |
|
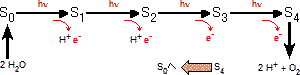
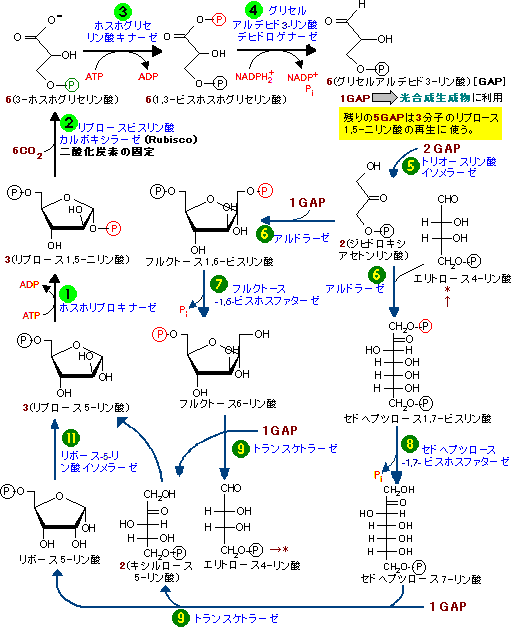
| 明反応において,光のエネルギーを利用してATPとNADPH2+を合成した。これらを用いて,二酸化炭素から糖を合成する過程を暗反応(dark reaction),Calvin サイクル,または還元的ペントースリン酸回路という。暗反応では光のエネルギーを一切必要としない。 暗反応は次の2つの段階に分けられる。以下の記述はサイクルが計3回まわった場合である。 |
|
| 1. | 還元的合成過程:段階 3分子のリブロース-5-リン酸と3分子のCO2から6分子のグリセルアルデヒド-3-リン酸(GAP)が合成される。この時,全部で9分子のATP(段階 |
| 2. | 再生過程:段階 1分子のGAPは糖の合成に使われる(光合成生成物)。残り5分子のGAPは糖の組み替えを経て,3分子のリブロース-5-リン酸に再生される。ここでは,自由エネルギーATPや還元剤NADPH2+を全く必要としない。再生過程はヘキソースリン酸側路(HMS,ホスホグルコン酸回路)と大変良く似ている。 |
| 以上,2段階をまとめると, 3 CO2 + 9 ATP + 6 NADPH2+→ GAP + 9 ADP + 8 Pi + 6 NADP+ となる。これはまさに,二酸化炭素を還元して糖(GAP)を創り出したことに他ならない! |
|
| グリセルアルデヒド-3-リン酸(GAP)から,糖新生やその他の経路によって,ショ糖,デンプン,セルロース,脂肪酸,アミノ酸などが合成される。 | |
|
|
 |
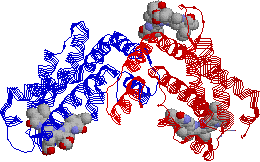
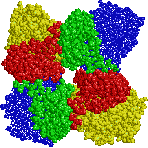 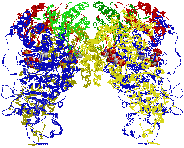 [ほうれん草Rubiscoの立体構造] (左)真上から見た図,(右)横から見た図 |
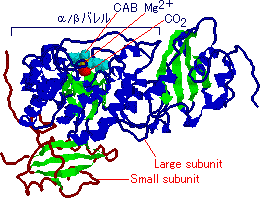
| [SynechococcusのRubiscoの大小サブユニット] a-らせん: 青,b-シート: 緑, CAB: 水色,基質(CO2): 赤 |
ほうれん草Rubiscoは大サブユニット8つと,小サブユニット8つから成る十六量体であるが,ここにはその半分の八量体が示されている。 酵素活性を抑えるために,2-カルボキシアラビニトール 1-リン酸(CAB)が結合した状態で結晶解析がされた。 |
| Synechococcusの酵素は大サブユニットと小サブユニットから成る十六量体タンパク質である。 大サブユニットにはトリオースリン酸イソメラーゼに見られるa/bバレルがある。 |
光呼吸 (Photorespiration)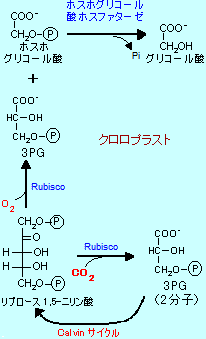 |
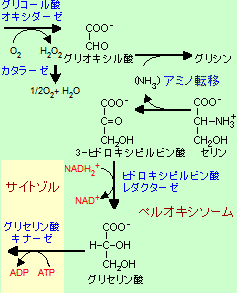 |
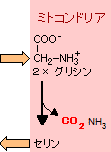 ペルオキシソームでは光呼吸 だけでなく,カタラーゼによる 過酸化水素の分解やグリオ キシル酸回路も行われる。 |
|
Rubiscoの阻害物質(CAB)
植物で夜だけつくられる2-カルボキシアラビニトール 1-リン酸 (CAB) は,CO2と結合した酵素(カルバモイルRubisco)に強く結合し,酵素活性を阻害する。これは光がない状態で光合成を抑える働きをしている。
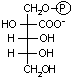 |
| [CABの構造式] |
C4経路による二酸化炭素の濃縮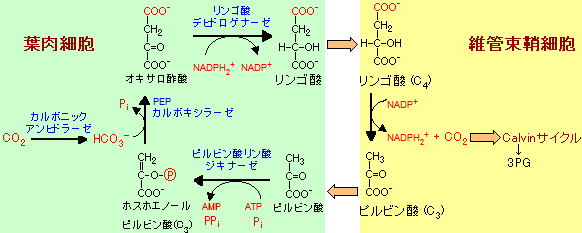
|
|