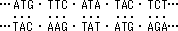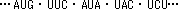|
|
 
|
核酸は次のように五炭糖、リン酸および塩基から成る。五炭糖にはD-リボースとデオキシ-D-リボースの2種があり、リボ核酸(RNA)にはD-リボース、デオキシリボ核酸(DNA)にはデオキシ-D-リボースが含まれる。塩基はアデニン(A)、グアニン(G)、シトシン(C)、ウラシル(U)、チミン(T)の5つがあるが、RNAではA, G, C, U、DNAではA, G, C, Tが含まれる。
| 核酸 | 五炭糖 | b-D-リボース | ・・・RNAにのみ存在 | 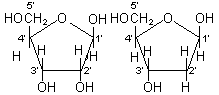 |
| 2-デオキシ- b-D-リボース |
・・・DNAにのみ存在 | |||
| リン酸 | H3PO4 | D-リボース デオキシ-D-リボース | ||
| 塩基 (base) |
プリン系 | ・・・アデニン、グアニン | ||
| ピリミジン系 | ・・・シトシン、ウラシル(RNAのみ)、チミン(DNAのみ) | |||
 |
 |
 |
 |
 |
 |
 |
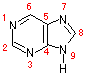
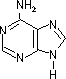
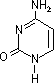
| プリン | アデニン (A) |
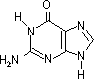
グアニン (G) |
ピリミジン | ウラシル (U) |
シトシン (C) |
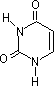
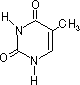
チミン (T) |
| [核酸を構成する塩基] | ||||||
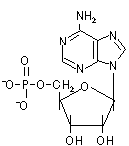
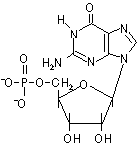
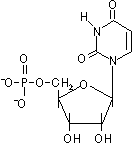
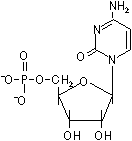
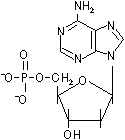
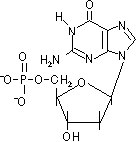
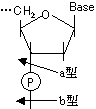
D-リボースやデオキシ-D-リボースの1'位に塩基が結合した化合物をヌクレオシド(nucleoside)という。ヌクレオシドの5'位にリン酸がエステル結合した化合物をヌクレオチド(nucleotide)という。
 |
 |
 |
 |
| アデノシン5'-リン酸 (AMP) |
グアノシン5'-リン酸 (GMP) |
ウリジン5'-リン酸 (UMP) |
シチジン5'-リン酸 (CMP) |
| [RNAを構成するリボヌクレオチド] | |||
 |
 |
 |
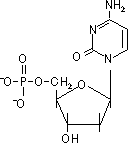 |
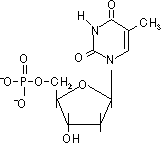
| デオキシアデノシン 一リン酸(dAMP) |
デオキシグアノシン 一リン酸(dGMP) |
デオキシチミジン 一リン酸(dTMP) |
デオキシシチジン 一リン酸(dCMP) |
| [DNAを構成するデオキシリボヌクレオチド] | |||
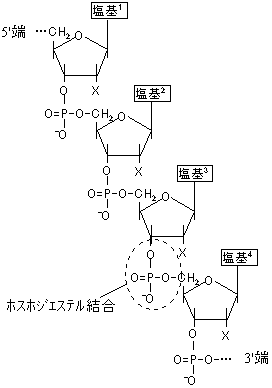
核酸(DNAやRNA)は、ヌクレオチド単位が長く連結した鎖状の高分子化合物である。
 |
各種DNA中のプリン,ピリミジンの含量と シャルガフ則
|
||||||||||||||
シャルガフ則 A/T=1、G/C=1、 プリン/ピリミジン=1 |
|||||||||||||||
| [核酸のポリヌクレオチド鎖] | |||||||||||||||
| X=H (DNA), OH (RNA) 塩基1-4=AGCT (DNA), AGCU (RNA) |
|||||||||||||||
|
|
 デオキシリボ核酸(DNA,deoxyribonucleic acid)
デオキシリボ核酸(DNA,deoxyribonucleic acid)
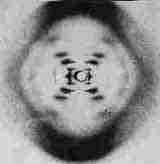 [DNAのX線回折像] |
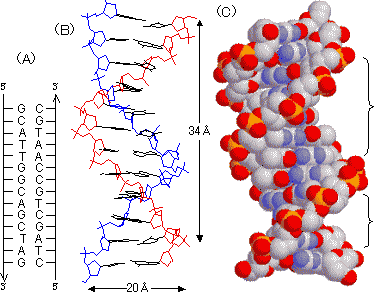 [DNAの二重らせん構造のモデル] |
主溝 副溝 |
| (A) DNA二本鎖間の塩基対: 2本の鎖は互いに逆向きである。 (B) 二重らせんの針金模型: らせんは右巻き。内側に向いた塩基対を構成するプリンとピリミジン環はらせん軸にほぼ垂直。 (C) 二重らせんの空間充填模型: らせんの側面には深い溝(主溝, major groove)と浅い溝(副溝, minor groove)がある。 |
||
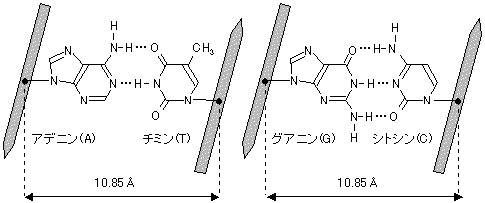
DNAの鎖は相補的な塩基対を形成し,互いに相手の鎖を認識できる。
[DNA中のチミン(T)とアデニン(A)、シトシン(C)とグアニン(G)間の水素結合 ]
Watson-Crick塩基対という。
A・・・・T 相補的塩基対(complementary base pair)
G・・・・C <分子生物学の基礎>
この原理を利用し、DNAのそれぞれのポリヌクレオチド鎖(親鎖)を鋳型として、それらに相補的な新しい鎖(娘鎖、daughter chain)が合成される。新しい鎖を構成する2本の鎖の一方は元の親鎖由来である。これを半保存的複製(semi-conservative replication)という。詳細についてはDNA複製を参照せよ。
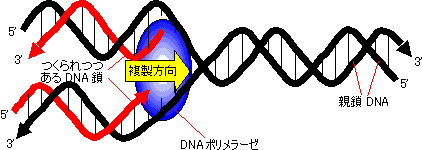
[DNAの半保存的複製のモデル] 赤い鎖が新しくつくられた鎖で、親鎖由来の黒い鎖と逆平行である点に注意。
リボ核酸(RNA、ribonucleic acid)
細胞質と核に存在。DNAの二本鎖のうち一方を鋳型として、A→ U,T→ A,G→ C,C→ Gの規則に従って合成される。
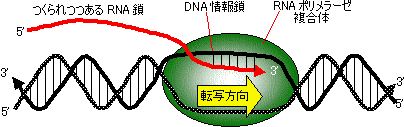
| [RNAの合成のモデル] |
| DNAの2本鎖のうち、鋳型となる鎖(黒線)は3'から5'方向に 読みとられ、5'から3'方向にRNA(赤い線)がつくられる。 |
タンパク質の生合成に関与する主なRNAとしては、次の3種類がある。
伝令RNA(messenger RNA,mRNA)
- DNAのタンパク質の1次構造に関する情報を写しとったもの。細胞質に移り,リボソームに結合してタンパク質合成の鋳型となる。
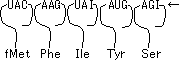
mRNAの3つ組塩基は1つのアミノ酸を指定する。連続した3つの塩基をコドン(codon)といい、1つのアミノ酸に対応する。
(例)GGG → Gly(グリシン)
AUGは開始コドン。下線は終止コドン。
mRNAのコドン表 2文字目 U C A G 1
文
字
目U UUU Phe
UUC Phe
UUA Leu
UUG LeuUCU Ser
UCC Ser
UCA Ser
UCG SerUAU Tyr
UAC Tyr
UAA オーカー
UAG アンバーUGU Cys
UGC Cys
UGA オパール
UGG TrpU
C
A
G3
文
字
目C CUU Leu
CUC Leu
CUA Leu
CUG LeuCCU Pro
CCC Pro
CCA Pro
CCG ProCAU His
CAC His
CAA Gln
CAG GlnCGU Arg
CGC Arg
CGA Arg
CGG ArgU
C
A
GA AUU Ile
AUC Ile
AUA Ile
AUG MetACU Thr
ACC Thr
ACA Thr
ACG ThrAAU Asn
AAC Asn
AAA Lys
AAG LysAGU Ser
AGC Ser
AGA Arg
AGG ArgU
C
A
GG GUU Val
GUC Val
GUA Val
GUG Val*GCU Ala
GCC Ala
GCA Ala
GCG AlaGAU Asp
GAC Asp
GAA Glu
GAG GluGGU Gly
GGC Gly
GGA Gly
GGG GlyU
C
A
G
* 原核生物では開始コドンとなる。
5'末端にキャップ構造,3'末端にポリA鎖をもつ。 タンパク質の1次構造に対応する部分をコード領域という。

- 細胞質中に存在する低分子量のRNA。A,G,U,C以外の特殊塩基が含まれる。3‘末端は--CCAで、ここにアミノ酸をエステル結合し、リボソームへと運ぶ。分子中の1ヶ所にアンチコドン(暗号解読部)部位があり、mRNAと結合する。tRNAはタンパク質と核酸の橋渡しをする分子である。
(例) コドン: 5'-A-G-A- 3' (mRNA)
アンチコドン: 3'-U-C-U- 5' (tRNA)- リボソームはタンパク質合成の場で、大腸菌では3種のrRNAと53種のタンパク質、真核細胞では4種のrRNAと82種のタンパク質から成る。リボソームの重量の2/3はrRNAが占めている。
|
|
アルカリに対する安定性
加水分解酵素
|
||||||||||||||||||||
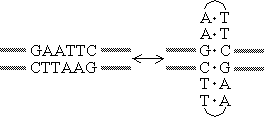 |
| [回文(パリンドローム palindrome)構造の例] 右のように、十字型の構造をとることができる。 |
核酸の変性とアニーリング
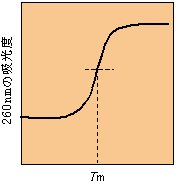 [DNAの融点] |
融点は溶媒、溶液中のイオンの種類と濃度、pH、G:C含量などで変わる。 A:Tは水素結合2本、G:Cは水素結合3本なので、G:C含量が高いほど融点も高い。 |
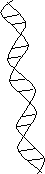 |
弱い加熱 |
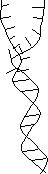 |
強い加熱 |
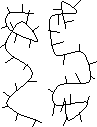 |
| 冷却 (アニーリング) |
冷却 | |||
| 2本鎖らせん構造 | 1本鎖ランダム構造 (変性状態) |
|||
| [DNAの変性とアニーリング] | ||||
|
|
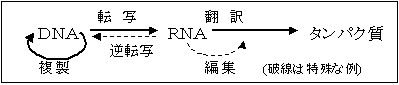
DNAからRNAがつくられ,RNAからタンパク質がつくられる。このような遺伝情報の流れを中心教義(セントラルドグマ)とよぶ。ただし,逆転写酵素の発見,RNAエディティングの発見により,この教義は多少の訂正を余儀なくされている。
DNA 複製
転写 アンチコドン
mRNA 翻訳 アミノ酸 タンパク質 [遺伝情報の流れ(DNAからタンパク質まで)]
遺伝情報に関する4つの基本過程 (1) 遺伝情報の保存……DNA複製 (DNA replication)
(2) 遺伝情報の維持……DNA修復 (DNA repair)
(3) 遺伝情報の発現……タンパク質合成 (転写と翻訳)
(4) 遺伝情報の改良……遺伝的組み替え
|
|