
|
細胞外マトリックス 細胞接着分子 接着分子の異常 |
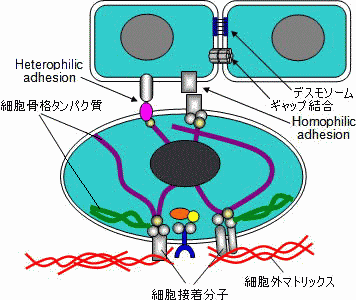 |
| 細胞間認識と細胞接着 |
|
|
|
| 細胞外マトリックス |  |
|
|
|
 細胞外マトリックスと細胞接着
細胞外マトリックスの種類
細胞外マトリックスと細胞接着
細胞外マトリックスの種類
 コラーゲン(collagen, COL)
コラーゲン(collagen, COL)
| 分類 | 分子種 | 特徴 |
|---|---|---|
| 1. フィブリル形成型 | I,II,III,V,XI | 7 nm周期の縞模様をもつフィブリルを形成。 |
| 2. ファシット型 | IX, XII, XIV, XVI, XIX | 三重ラセンが中断された線維会合型。 |
| 3. 短鎖型 | VIII, X | シートを形成。 |
| 4. 基底膜型 | IV | 全ての基底幕の成分。 |
| 5. マルチプレキシン | XV, XVIII | 三重ラセンが何度も中断された構造。 |
| 6. マシット型 | XIII, XVI | 細胞表面分子型。細胞接着分子? |
| 7. その他の型 | VI VII |
じゅず状フィラメント(ミクロフィブリル)を形成。 足場となる繊維(アンカリングフィブリル)を形成。 |
 |
| ウシtypr I (a1) コラーゲンのアミノ酸配列 |
| 黄色の背景のGは三連配列の1文字目のGlyを示す。この領域はポリプロリンII型の三重ヘリックス構造をとる。この構造でProのC=OとGlyのN-H間に水素結合が生じる。また,このGly残基が他の残基に変わると三重ヘリックス構造は変形する。 |
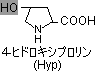  |
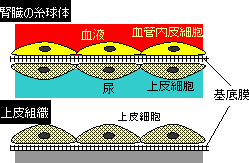 |
| コラーゲンの三重ラセン構造 | 基底膜 |
フィブロネクチン(fibronectin, FN)
 |
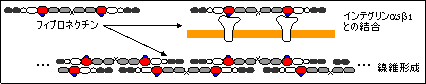 |
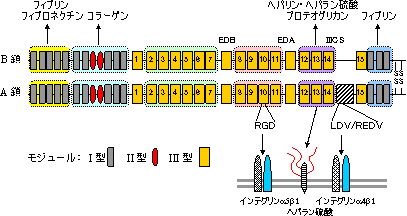
| フィブロネクチンの構造 |
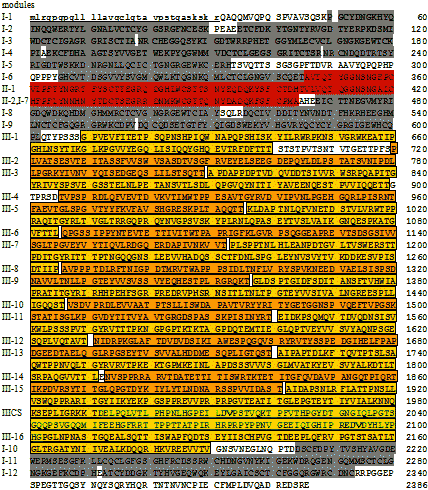 |
| フィブロネクチン前駆体の1次構造 |
| 小文字下線部,シグナルペプチド; 灰色,モジュールI型; 赤,モジュールII型; 橙と黄色,モジュールIII型 52-272: フィブリン-ヘパリン結合部位。 308-608: コラーゲン結合部位。 2206-2337: フィブリン結合部位 464-477: コラーゲン結合に必須。 太字: 細胞接着部位(RGD/PHSRN/LDV/REDV)。 |
| 【モジュールとは】 タンパク質の1次構造で,ある特徴をもつ繰り返し配列。しばしば他のタンパク質にも 同じモジュールが見出される。 |
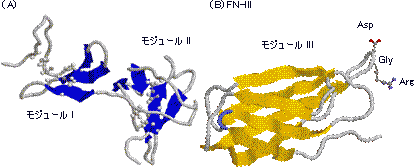 |
| フィブロネクチンの3つのモジュールの立体構造 |
| (A)コラーゲン結合モジュールI型とII型。それぞれ,2つのS-S結合がある。 (B)モジュールIII。S-S結合はない。右上のループにRGD配列が存在。 |
ラミニン(laminin, LM)
 |
|
||||||||||||||||||||||||
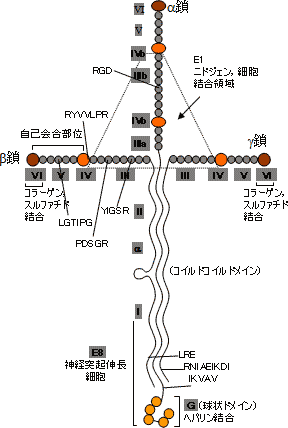
| ラミニンの構造モデル | |||||||||||||||||||||||||
| a鎖,b鎖,g鎖の3つのサブユニットから成る十字架形。 球状ドメインが細胞接着部位。 I 〜VI , G , E8 , a はドメイン名。 アミノ酸配列は種々の機能に関与するもの。 |
|||||||||||||||||||||||||
フィブリノーゲン
 |
| フィブリノーゲンの三重ラセン構造 |
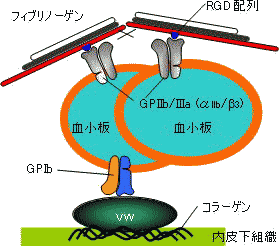 |
| 血小板凝集とフィブリノーゲン |
| GPIIb/IIIa,フィブリノーゲンを結合するインテグリン; vW,フォンビルブランド因子; GPIb,vW受容体となるインテグリン. |
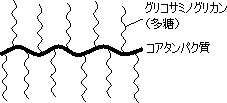
プロテオグリカン
 |
 |
 |
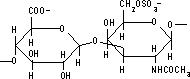 |
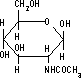
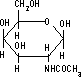
| プロテオグリカンの構造 | N-アセチル-D-グルコサミン | N-アセチル-D-ガラクトサミン | コンドロイチン 6-硫酸 |
|
グリコサミノグリカン鎖としては,HA, CS, DS, HS, KS, ヘパリンなど。 二糖の構成単糖にはN-アセチル-D-グルコサミンかN-アセチル-D-ガラクトサミンが含まれる。 二糖の相手としては,D-グルクロン酸,L-イズロン酸, D-ガラクトースなど。結合位置は3または4位。 |
|||
|
|
|
| 細胞接着分子 |  |
|
|
|
細胞膜には多くの膜タンパク質(接着分子)が存在し,細胞間の相互作用に関与。
細胞接着分子の種類
カドヘリンスーパーファミリー クラシックカドヘリン(cadherin)
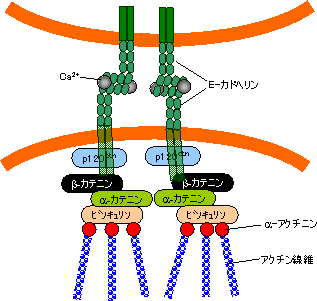 |
| Eカドヘリンを介した細胞接着 |
ノンクラシックカドヘリン
インテグリンファミリー
|
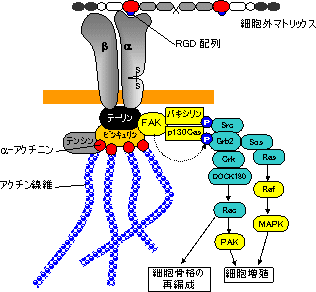 インテグリンを介するシグナル伝達 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| COL, コラーゲン: LM, ラミニン: FN, フィブロネクチン: FB, フィブリノーゲン: VW, フォンビルブラント因子: VN, ビトロネクチン: TSP, トロンボスポンジン: OS, オステオポンチン。―は結合配列が不明。 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
セレクチンファミリー
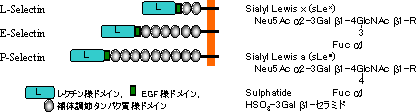 |
| セレクチンファミリーと結合糖鎖の構造 |
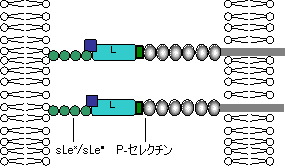 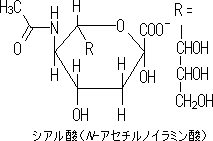 |
| セレクチンと糖鎖リガンドの接着を示す模式図 |
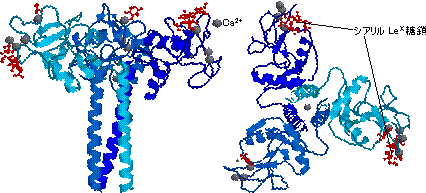 |
| セレクチンのレクチン様ドメインとシアリルLex糖鎖の相互作用 |
| 左は横から見た図,右は真上から見た図。灰色の丸はCa2+ |
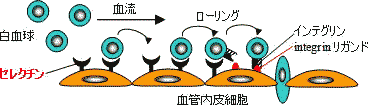 |
| 炎症部位への白血球の遊走と細胞接着分子 |
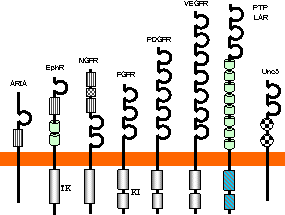
免疫グロブリンスーパーファミリー
免疫グロブリン(Ig)スーパーファミリーの仲間

 |
| 免疫グロブリン(Ig)ドメイン |
| 7本の逆並行b-シートで筒状構造を形成。S-S結合が少なくとも1つある(左図を参照)。 |
免疫グロブリンスーパーファミリーのドメイン構造
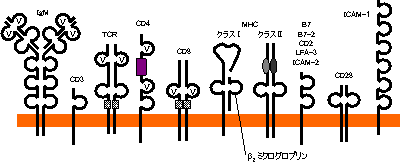
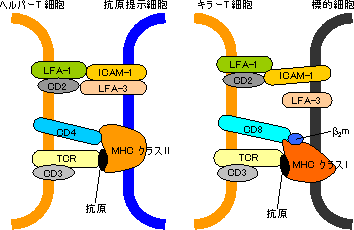 |
| 免疫グロブリンファミリータンパク質の機能の例 |
| (左)ヘルパーT細胞と抗原提示細胞の相互作用, (右)キラーT細胞と標的細胞の相互作用 |
| (単純型) | (複合型) |
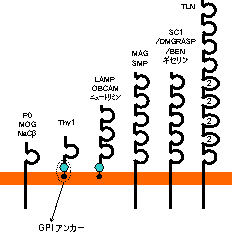 |
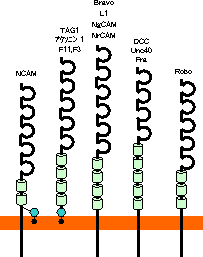 |
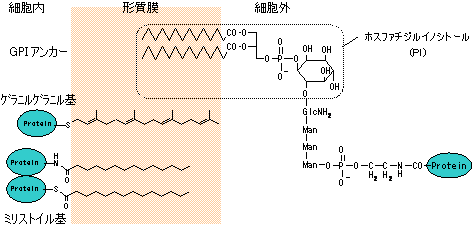 |
| 真核細胞の細胞膜へのタンパク質のアンカリング |
| GPI: グリコシルホスファチジルイノシトール |
|
|
|
| 接着分子の異常 | |
|
|
|
臓器拒絶: ICAM-1とLFA-1,VCAM-1とVLA-4,CD28とB7。ウィルス感染における標的: CD4(HIV)。MHCクラスⅠ(サイトメガロウィルス)。ICAM-1(ライノウィルス)。膜のシアル酸(センダイウィルスやインフルエンザウィルス)。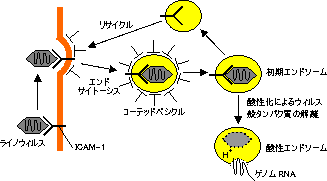 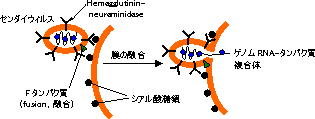 |
| ウィルス感染における細胞膜成分の関与 |
| (A) ICAM-1はライノウィルスの標的。 (B) センダイウィルスは膜シアル酸に結合し,Fタンパク質の作用で 宿主細胞膜と融合。 |
慢性関節リウマチ: インテグリンファミリー(LFA-1, VLA-4, VLA-1),Igスーパーファミリー(ICAM-1,VCAM-1),セレクチンファミリー(E-セレクチン)などが関与。呼吸器疾患: LECAM,CD18とICAM-1。ガンの転移: セレクチンファミリー,カドヘリンファミリー。白血球粘着異常症: インテグリンb2の欠損→ローリングの異常。
|
|