|
|
 
|
|
|
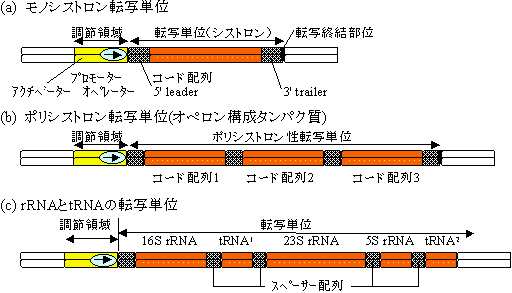
《ヒトのゲノムDNAには何が書かれているのか?》
| 遺伝子 (30%) | 非遺伝子 (70%) | ||||
| 3 | 非コード領域 (27%) | 偽遺伝子とスペーサー (52%) | (18%) | ||
| コード領域 (3%) | 繰り返し配列 | ||||
|
1) 遺伝子領域: ゲノム全体の30%(900Mb) |
タンパク質とrRNA,tRNAをコードする約22,000個の遺伝子があるとされている。
 構造遺伝子領域(90Mb=全体の3%)
構造遺伝子領域(90Mb=全体の3%)
 非コード領域(810Mb=全体の27%)
非コード領域(810Mb=全体の27%)
| 遺伝子産物 | 生物 | エキソン | イントロン | |
| 数 | 総塩基対 | |||
| エリトロポエチン a-インターフェロン アデノシンデアミナーゼ 低密度リポタンパク質 チログロビン VIII因子 アポリポタンパク質 b-グロビン HGPRT ビテロゲニン フィブロイン(絹) ファセオリン ウリカーゼサブユニット ゼイン tRNATyr シトクロムb |
ヒト ヒト ヒト ヒト ヒト ヒト ヒト マウス マウス カエル カイコ インゲンマメ ソラマメ トウモロコシ 酵母 酵母ミトコンドリア |
582 600 1,500 5,100 8,500 9,000 14,000 432 1,307 6,300 18,000 1,263 300 700 76 2,200 |
4 0 11 17 >40 25 28 2 8 33 1 5 7 0 1 6 |
1,562 0 30,000 40,000 100,000 177,000 29,000 762 32,000 20,000 970 515 4,500 0 14 5,100 |
植物,動物,それらに感染するウイルスにはイントロンが多いが,無脊椎動物ではまれ。原核生物にはないが,T4バクテリオファージのチミジル酸シンテターゼにはイントロンがある。古細菌にもイントロンが見つかっている。
|
2) 非遺伝子領域: ゲノム全体の70%(2100Mb) |
機能不明ものが多い![]() 従来,ガラクタ(junk)DNAといわれてきた。最近,マウスゲノムの発現の解析から、全体の長さの60%以上が何らかの形で細胞内で利用されていることが報告された(2005年)。人でも同様の仕組みがあると考えられるので、下記の非繰り返し配列にはまだ機能未知のものが含まれる可能性が高い。
従来,ガラクタ(junk)DNAといわれてきた。最近,マウスゲノムの発現の解析から、全体の長さの60%以上が何らかの形で細胞内で利用されていることが報告された(2005年)。人でも同様の仕組みがあると考えられるので、下記の非繰り返し配列にはまだ機能未知のものが含まれる可能性が高い。
非繰り返し配列(1680Mb=ゲノム全体の52%)
繰り返し配列(420Mb=全体の18%)
《Alu配列》
制限酵素Alu Iで切断される部位を持つ全長300bpの配列で、
ゲノム全体にほぼ均一に存在。
|
|
ミトコンドリアの構築や機能に必要なタンパク質の大部分は核DNAの遺伝子でつくられるが、一部はミトコンドリアゲノムにコードされている。ゲノムの大きさや遺伝子の構成は種によって大きく異なる。動物では20kbp以下、植物では数100〜数1000kbp、酵母は約80kbp。1つのミトコンドリアに5〜10コピー存在する。
【ヒトミトコンドリアDNAの特徴】
全体的には原核生物的な特徴が見られるが,mRNAがポリA化されるなど真核生物的性質も兼ね備えている。
ゲノムの構造
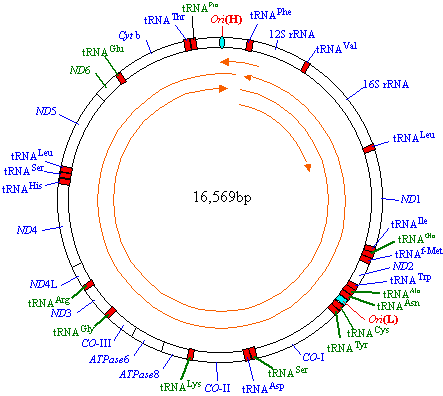 |
| [ヒトミトコンドリアDNA] |
| DNA2本鎖(H鎖,L鎖と呼ぶ)のそれぞれに複製起点(OriHとOriL)が存在。 内側の矢印はOriHからの転写産物。一番内側の矢印はrRNA専用の転写。 OriLからは全長のものは1つだけつくられる。 遺伝子の向きは時計回りのものと,反時計回りのものとがある。 赤いboxはtRNA遺伝子。 ND, NADHデヒドロゲナーゼ複合体; CO, シトクロムcオキシダーゼ;Cytb, シトクロムb |
ミトコンドリアの分裂とDNA複製
転写
翻訳
|
|
遺伝子(gene)は、個々の表現形質に対応する原因となる単位を表わし,分子的には独立した存在ではなく,単に巨大なDNAの一部である。これに対して、ゲノム(genome)という概念は、ある生物種の個体全体を完全な状態に保つために必要な遺伝的情報の1セットを指す言葉である。従って,生物種ごとに固有のゲノムが存在する。ウイルスや細菌についてもファージゲノムや細菌ゲノムとよぶ。細胞小器官でDNAをもつミトコンドリアや葉緑体についても、ミトコンドリアゲノム、葉緑体ゲノムと呼ぶことができる。遺伝情報そのものはDNAの塩基配列であるから,ゲノムの実体はDNAそのものといえる。
今後、たくさんの生物のゲノムの全構造が決められ、 その中の遺伝子と遺伝子産物に関する情報が次々と明らかにされるであろう。ゲノム全体を対象に全遺伝情報を調べあげ、そこから個別的な研究を始めるという研究を「ゲノム生物学」という。
【全ゲノム配列が発表・公開された生物】
バクテリアを中心に、ゲノムの全配列が次々と報告されています。これまでに全ゲノム配列が 決定されているものは以下のとおりである(from KEGG)。DNAの大きさは生物種ごとにずいぶん異なる。現在のところ,真核生物で全ゲノムが分かっているのはC. elegans(線虫)だけであるが,2003年までにはヒトゲノムの全配列が明らかとなるであろう。
| 生物種 | ゲノムサイズ (kbp) |
遺伝子総数 |
| Mycoplasma genitalium (マイコプラズマ感染症) Mycoplasma pneumoniae (肺炎の病原体) Borrelia burgdorferi B31(ライム病の病原スピロヘータ) Chlamydia trachomatis (トラコーマ病原体) Rickettsia prowazekii (発疹チフス病原体) Treponema pallidum (梅毒トレポネマ) Chlamydia pneumoniae (クラミジア) Aquifex aeolicus(超好熱性細菌) Helicobacter pylori J99(ピロリ菌) Methanococcus jannaschii (メタン細菌) Helicobacter pylori 26695(胃炎や胃潰瘍の病原菌ピロリ菌) Pyrococcus horikoshii OT3(超好熱性古細菌) Methanobacterium thermoautotrophicum deltaH(メタン細菌) Haemophilus influenzae Rd(インフルエンザ菌) Archaeoglobus fulgidus (好熱性硫黄細菌) Synecocystis sp.PCC6803 (ラン藻類) Bacillus subtilis (枯草菌) Mycobacterium tuberculosis H37Rv (結核菌) Escherichia coli (大腸菌) Saccharomyces cerevisiae (出芽酵母) Caenorhabditis elegans (線虫) Thermotoga maritima MSB8 (好熱性真正細菌) Aeropyrum pernix K1 (好気性好熱菌) |
580 816 910 1040 1110 1140 1130 1550 1640 1660 1670 1740 1750 1830 2180 3570 4210 4410 4640 12000 97000 1860 1670 |
467 677 1256 894 834 1031 - 1522 1495 1770 1566 1979 1869 1717 2407 3166 4100 3918 4289 6286 約19000 1877 2694 |
|
|